成都生物所齐银团队揭示沙蜥两性颜色变异的遗传因素和进化机制
MBE | 沙蜥两性颜色变异的遗传因素和进化机制

Bin Lu, Xia Qiu, Weizhao Yang, Zhongyi Yao, Xiaofeng Ma, Shunyan Deng, Qi Zhang, Jinzhong Fu, Yin Qi, Genetic basis and evolutionary forces of sexually dimorphic colour variation in a toad-headed agamid lizard, Molecular Biology and Evolution, 2024;, msae054,
动物的体色在适应和社会交流中发挥着至关重要的作用。由于生活环境和社会交流环境的复杂性,动物经常表现出颜色变化,这可能有助于性别识别、个体身份以及信号质量或行为策略。蜥蜴的特点是颜色多种多样,是色彩最鲜艳的脊椎动物之一。这种显著的变化在物种和种群内部和种群之间以及跨性别之间都很明显,强调了蜥蜴与其复杂的生态位之间的动态相互作用。蜥蜴的颜色是一种多方面的性状,受到多种因素的影响,包括栖息地适应、反捕食策略(例如伪装或躲避捕食者)和社会互动(为了吸引配偶或保护资源)。这些选择压力的相互作用很可能塑造了当今不同蜥蜴种群中观察到的显著颜色变化。了解这种变异的遗传基础,因为蜥蜴的颜色在它们的生存和繁殖中起着关键作用,可以为这些迷人生物的生态和进化提供有价值的见解。通过深入研究控制颜色图案的遗传机制,研究人员可以更深入地了解蜥蜴与其环境之间的复杂关系。
贵德沙蜥(Phrynocephalus putjatai)为解开蜥蜴两性二态性颜色的遗传基础提供了一个理想的系统。值得注意的是,它缺乏异形性染色体,并且其身体颜色在不同地理区域表现出差异。具体来说,贵南县木格滩沙漠中的种群(以下统称为P. guinanensis,GN)表现出惊人的性别二态性和与年龄相关的颜色变化。成年雄性呈现红色,而未成年和成年雌性则呈现蓝色,主要是在身体侧面(图 1a)。相比之下,贵德县和附近地区(以下简称 GD)的种群及其近亲青海沙蜥(例如P. vlangalii)通常表现出柔和的颜色。鉴于它们的近亲分布以及关于 GN 和 GD 物种分类的长期争论,归因于它们相似的遗传背景,研究人员假设 GN 的颜色变化可能是最近通过有限数量的基因改变而出现的。此外,分布在位于木格滩沙漠和贵德县之间的过马营镇(以下简称GM)的种群表现出中间表型(图1a),暗示GN和GD种群之间可能存在基因渗入。为了阐明控制颜色变化形成和维持的过程,将GM种群纳入分析中。
这项研究的主要目的是阐明 GN 群体中性别特异性颜色变异的遗传基础,并揭示导致与颜色相关的基因固定的潜在进化力量。方法涉及多个方面的全面分析。最初,量化了不同群体和性别之间的颜色变化。随后,深入研究了色素沉积差异背后的生化和细胞基础。为了探索颜色变异的遗传机制,研究人员对三个沙蜥种群(GN、GD、P. vlangalii)进行了全基因组测序。利用 GN 染色体水平组装作为参考,使用群体基因组学对来自不同颜色的 100 个个体进行了重测序。最后,将现场实验与视觉建模技术相结合,以仔细研究影响 GN 种群颜色变化演化的可能存在的选择压力。这种全方位的方法旨在全面了解导致沙蜴颜色变化的遗传、生化和生态因素。

Graphical Abstract颜色变化的表型、细胞和生化基础
为了验证 GD、GM 和 GN 群体之间明显的颜色差异,以及 GN 群体中的性别二态性颜色,利用光谱测定法来量化体色。共检测了来自贵德县的GD1和GD2、贵南县的GN1和GN2以及过马营镇的GM1和GM2等6个群体的268个体(图1a)。基于色调(峰值反射率的波长)和色度(峰值的波长带宽)的光谱曲线(图1b)、聚类分析(图1c和补充图1)和线性模型反射率共同证实了 GD、GM 和 GN 种群个体之间的颜色差异,以及 GN 种群中存在性别二态性颜色。由于颜色变化明显,将GD、GM和GN分为不同的颜色组,以便于后续比较。

Fig. 1 | Colour variation in Phrynocephalus putjatai.

Fig. S1 Cluster analysis based on hue and chroma.
为了阐明颜色变化的生化基础,采用液相色谱质谱法 (LC-MS) 来量化 GD 和 GN 群体中两性表皮中的类胡萝卜素和蝶啶含量。结果显示,不同肤色对应的特定代谢物浓度不同(图 1f)。红色表皮样本表现出低水平的黄色色素,包括蝶啶黄蝶啶、类胡萝卜素-隐黄质、-胡萝卜素和叶黄素(图1f)。与 GD 相比,GN 表皮中-胡萝卜素和黄蝶啶的浓度显著较低,尤其是 GN 雄性的-胡萝卜素(图1f)。两种无色蝶啶色素,6-生物蝶啶和异黄蝶啶,可能作为有色色素的前体,与蓝色和暗色表皮相比,在红色表皮中含量更丰富(图 1f)。因此,红色、蓝色和暗色表皮的区别可能源于不同浓度的黄色类胡萝卜素和彩色蝶啶。然而,存在显示色素含量不显著差异的实例,特别是在 GN 雄性和雌性之间,可能归因于一些个体异常值和相对较小的样本量。一般来说,与蓝色和暗色表皮相比,红色表皮表现出相对较低浓度的类胡萝卜素和相对较高浓度的蝶啶。
研究结果提供了对蜥蜴种群颜色变化的结构和生化基础的见解,强调了类胡萝卜素和蝶啶在形成观察到的差异中的作用。
染色体水平基因组构建和系统发育分析
作为研究蟾蜍蜥蜴颜色变异遗传基础的资源,使用从单个雌性 GN 蜥蜴中提取的 DNA 衍生的长读长序列生成了染色体水平的参考基因组。利用 Hi-C 方法并在已建立的核型研究(2n = 48)的基础上,GN 的contig被挂载到 24 对染色体,产生总大小为 1.81 Gb 的基因组,与估计的基因组大小匹配 (1.83 Gb) 。该组装件的scaffold N50 为 132.7 Mb,L50 为 6,间隙为 3.4%(补充图 3)。为了进行比较分析,还使用 PacBio 长读长测序(80×~105×覆盖率)对 GD 蜥蜴和P. vlangalii的基因组进行了测序和组装。观察到至少 91% 的长读和 98.08% 的短读长比对率,脊椎动物单拷贝基因的完整性为 91.4%-92.2%(补充图 4),证实了 基因组组装的完整性、准确性和完整性。染色体共线性分析表明蜥蜴染色体起源于安乐蜥染色体的严重分离事件(补充图5)

Fig. S3 Genome landscape and synteny across GN, GD and P. vlangalii, showing regional variation (500-kb windows) in GC content and gene density.

Fig. S4 BUSCO assessment of the genome assembly integrity for GN, GD and P. vlangalii.

Fig. S5 Chromosomal synteny comparison between Anolis carolinensis and GN lizard.
系统发育分析(图2a)揭示了GN和GD之间的姐妹群关系。GN 和 GD 之间的估计分化时间约为 270 万年前(mya;图 2a)。该估计来自多个校准点,并与上新世末期一致。P. vlangalii与 GN 和 GD 的共同祖先在 6.95 mya 左右分化。值得注意的是,与其他有鳞类物种相比,亚洲蜥蜴进化枝表现出相对较高的中性进化替代率(每百万年每个位点约次替换)。有趣的是,颜色鲜艳的 GN 蜥蜴表现出化学感知相关基因的明显减少,包括嗅觉受体 (OR) 和犁鼻受体 2 型 (V2R) 家族(补充图6和7)。这一发现促使人们对视觉信号和化学感应适应之间潜在的相互作用进行研究。

Fig. 2 | Genomic landscape of divergence and demography.
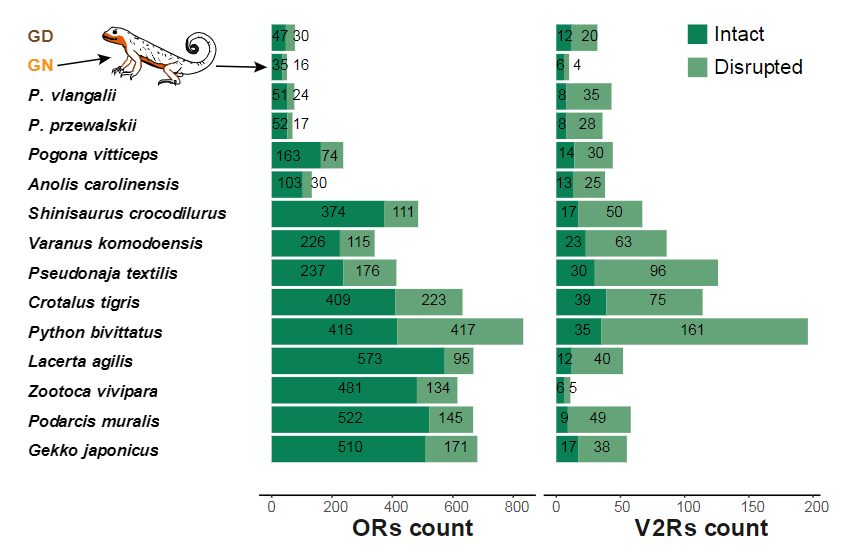
Fig. S6 The gene families associated with chemoreception, including olfactory receptor (OR) and vomeronasal receptor type-2 (V2R) genes, exhibit a contraction in GN lizard.

Fig. S7 Contraction of the chemoreception-related gene families in toad-headed agamid lizards. a Olfactory receptors tree. b Vomeronasal receptors type-2 tree.三个不同颜色群体的遗传组分和进化模拟
为了为揭示颜色变化的遗传基础,深入研究了三种不同颜色组(GN、GD 和 GM)的遗传组成和进化历史。利用来自 100 个个体的群体基因组数据,平均测序深度约为 11×,采用主成分分析 (PCA) 和 ADMIXTURE来描绘与每个颜色组相对应的清晰遗传簇(图 2b和c)。值得注意的是,观察到一定程度的遗传混合,特别是在GM和其他两组之间(图2c),这表明最近共享祖先、渐渗杂交或两者组合的潜在情况。基于个体的最大似然(ML)树进一步阐明了这种关系,揭示了来自GM的个体在GN进化枝的基部形成了一个并系群(图2d)。这种模式与 GM 群体中 GN 遗传成分的较高比例相一致,如 ADMIXTURE 分析所证明的那样。这种有趣的模式促使人们进一步探索这些群体的进化动态。
观察PSMC结果图,GN和GD群体的有效群体大小()的曲线具有一致的趋势(图2e)。自从上新世末期最初分化以来,这两个群体都经历了的增加,在早更新世末期达到峰值(~0.8 mya),随后经历持续下降, 末次冰期(LGP)早期阶段有暂时恢复。

Fig. S8 Schematic representation of 11 alternative demographic scenarios modeled, representing four possible scenarios with three gene flow patterns (no gene flow, recent gene flow and continuous gene flow).
溯祖模拟包含 11 种替代进化情景(补充图 8),表明二次接触是最合适的模型(模型 2)。这个模型表明了一种古老的孤立和最近形成的混合模式,具有西南到东北的迁徙路线。GN和GD在没有基因流的情况下在2.6 mya左右发生分化,与分子钟估计(~2.7 mya)紧密一致,后来在大约0.3 mya杂交产生GM(图2f)。混合参数()估计表明,大约80%的GM基因组起源于GN,而只有大约20%来自GD(图2f)。基于 ADMIXTURE 结果也证实了贡献的不平等(图 2c)。此外,自 GD 和 GN 之间的初始杂交事件以来,存在连续且不对称的基因流,主要迁移方向是从 GN 到 GD。与反方向(和)相比,从 GN 到GD() 和GM() 的基因流速率更高表明了这一点;图2f)。考虑到GN的有限分布、上新世末起源及其最初的隔离且没有迁移模式,有证据表明GN的生境可能出现在至少2.6mya,主要对应于青海高原隆升的第二阶段。
进化分析揭示了这些不同颜色群体错综复杂的遗传关系和进化历史,为更深入地了解这些种群颜色变化的遗传基础奠定了基础。
选择和候选位点的基因组信号
全基因组比较揭示了有限数量的强分化位点,其中 PBE 值大于 1 的顶级异常值主要聚集在五个小的基因组区域内(图 3a)。其中一个基因在 14 号染色体上跨越 80 kb,仅包含两个色素沉积基因:一个-胡萝卜素加氧酶 2 (BCO2) 基因和三个拷贝的 6-丙酮酰四氢生物蝶呤合酶 (PTS)。
BCO2基因与类胡萝卜素合成相关,与鸟类、哺乳动物和鱼类的非黑色素着色有关。鉴于检测到 GN 中的类胡萝卜素和蝶呤色素与 GD 群体相比存在不同水平,BCO2和PTS成为 GN 成熟雄性红色表皮的候选基因。有趣的是,发现了一个与攻击行为相关的高度分化的基因KIRREL3,位于 BCO2-PTS 位点附近,这意味着 GN 蜥蜴的攻击和颜色存在潜在的协同进化模式。
使用 PBS、FST 的成对比较和复合似然比 (XP-CLR) 检验进一步验证了五个局部 PBE 峰,但 GD-GM 对除外(补充图 9)。值得注意的是,前 1% XP-CLR 值显示出强烈的选择信号,特别是对于BCO2、PTS、IVL、MAP7、ATP2B2、SLC6A11、TAS2R41、DDX60、KIRREL3、SLC1A2等 PBE 候选基因。将颜色测量数据(色调、彩度和消色差)与群体序列数据结合起来,还进行了表型-SNP 关联分析 (GWAS)。然而,在 BCO2-PTS 基因周围没有检测到显著的 GWAS 信号。这可能归因于非遗传因素在决定颜色呈现方面发挥作用的可能性。除了遗传因素外,颜色定量还可能受到发育阶段(年龄)和摄入条件(因为类胡萝卜素必须通过饮食获得)的影响。这些因素可能会在表型测量中引入潜在的偏差,导致关联失败。
研究人员采用多方面的方法来识别和表征与颜色差异和其他性状相关的基因组区域,揭示蜥蜴与色素沉积、行为和视觉处理的潜在相关性。

Fig. 3 | Genomic basis for colour variation.性二态性颜色变异候选位点的特征
为了阐明潜在的遗传动力学,进一步对 BCO2-PTS 和MAP7位点内的分化区域进行了详细分析。有趣的是,两对在 BCO2-PTS 基因座内都表现出高度分歧的区域,值范围为 0.6 至 0.8(图 3b)。相反,GD 和 GM 对在同一区域显示出连续的低值 (< 0.2)。值得注意的是,GN 与其他群体之间的差异加剧主要发生在基因区域内,在基因间区域急剧减少。这意味着 GN 群体中BCO2和 PTS 基因区域的重组率降低,选择信号强。BCO2-PTS 基因座的核苷酸多样性 () 显著低于 14 号染色体和整个基因组的平均值,与强选择事件的预期影响一致。特别引人注目的是,与 GD 和 GM 相比,GN 中从BCO28号外显子到 BCO2-PTS 基因末端的核苷酸多样性明显减少(图 3b)。尽管 GN 显示出最低水平的全基因组连锁不平衡 (LD),但发现 GN 中的 BCO2-PTS 和MAP7位点的 LD 高于 GD 和 GM。这一发现进一步支持了这些目标候选基因经历了正选择的假设(图3c)。
在BCO2-PTS基因区域中,大多数差异(> 90%;820个SNP中的766个)在内含子区域内被鉴定到,而编码区仅表现出14个同义和5个非同义(错义)取代(图3f)。值得注意的是,在 BCO2 的8号外显子中观察到的 G 到 A 的转变(chr14:26,069,289;下文为 p26069289)导致残基处发生取代,将其从带负电的天冬氨酸变为极性、中性的天冬酰胺(D420N;图 3g)。这种取代是 GN 蜥蜴所独有的,该位点的 D 残基在其他有鳞动物中高度保守(图 3g)。其他有鳞动物的基因组主要以第一个密码子位置的 G 等位基因为特征,除 GN 外,没有其他 A 突变被表征,这意味着 G 是祖先等位基因,A 是衍生突变。在检查 100 个个体中的 p26069289 变异后,观察到突变纯合子 (AA) 主要 (~96%) 在 GN 群体中发现,而野生型纯合基因型 (GG) 主要 (94%) 局限于 GD和GM群体(图3h)。
为了探讨 GN BCO2 蛋白中 D420N 取代的功能含义,对 BCO2-类胡萝卜素进行了 3D 受体-配体对接分析。天冬酰胺 (N) 等极性氨基酸常见于蛋白质表面,在酶的底物识别和催化中发挥着至关重要的作用。构建的 3D 对接系统显示,420号残基与周围的四个残基一起形成埋在受体-配体界面处的结合域内的 α 螺旋二级结构。此外,胡萝卜素配体通过氢键(距离< 4 Å的氢键)与BCO2的420号残基相互作用(图4)。

Fig. 4 | Predicted three-dimensional structure model of BCO2--carotene complex.GN 中颜色变化的驱动力
鉴于在 GN 与邻近种群之间的 BCO2-PTS 和MAP7区域观察到的有限基因流,作者试图了解 GN 个体颜色变化背后的驱动力。假设与邻近地区(GM 和 GD)相比,在 GN 中观察到的颜色可能赋予其沙漠栖息地特定的优势。
为了探索自然选择的作用,研究人员采用视觉模型来评估GN雄性和雌性的颜色与深色土壤相比是否可以在沙丘中产生伪装效果,从而潜在地提高存活率(图5a)。在 GN 和 GM 地区,人们经常看到潜在的捕食者(补充图 14)。如果雄性和雌性的颜色在沙丘中具有很高的伪装效果,作者的假设是,与GM的典型栖息地深色土壤相比,它们的颜色在沙丘中可能不那么明显。研究结果表明,与深色土壤相比,沙丘中 GN 蜥蜴的显眼程度降低。具体来说,彩色对比度表现出下降(图 5b),消色差对比度也下降(图 5c)。这表明雄性和雌性 GN 蜥蜴都可能很好地融入沙丘,可能比生活在深色土壤中的蜥蜴具有更低的捕食率。这可能会阻碍 GN 蜥蜴离开沙地,并可能有助于促进颜色相关基因的固定。

Fig. S14 Colouration and predator avoidance.
鉴于明显的性别颜色二态性,作者进一步研究了颜色对雄性配偶选择和雄性竞争的影响,以阐明性选择的作用。假设认为,雌性蓝色的显眼程度可能是个体品质的一个指标,雄性可能会表现出对更亮蓝色的雌性的偏好。为了检验这一假设,进行了三组二分法雄性择偶试验:(1) 蓝光阻挡和亮度降低与对照(检查蓝色和亮度在配偶选择中的重要性),(2) 亮度降低与对照(研究亮度(而不是颜色)在雄性选择中是否至关重要),以及(3)蓝光阻挡和亮度降低与亮度降低(评估蓝色而不是亮度在雄性选择中的重要性)。所有试验均在一个选定的场地内进行,该场地由两个雌性的房间组成,位于现场的一名常驻雄性前面。这些隔室由透光率接近 100% 的玻璃制成(补充图 15;图 5d)。

Fig. 5 | Natural and sexual selection for the evolution of colour variation in GN lizards.

Fig. S15 The transmission spectra of the two types of filters, and a control treatment.
最初研究了蓝色和亮度对雄性择偶的影响。雄性可以在 MW(蓝色阻挡和亮度降低)雌性和 FULL(未过滤)雌性之间进行选择。与 MW 相比,雄性明显更喜欢 FULL 雌性(图 5e),这表明蓝色和亮度可能是择偶决策中的重要因素。
实验旨在了解 GN 种群颜色变化的生态和进化驱动因素。这些结果支持了这样的假设:GN 个体的颜色可能受到自然选择和性选择的共同影响,在沙漠栖息地具有潜在的适应优势,并对配偶选择和雄性之间的竞争动态产生有意义的影响。
结论
通过整合颜色表型、表皮超微结构、色素生化、基因组和转录组数据以及现场行为实验,研究人员对蟾蜍蜥蜴两性二态性颜色变异背后的遗传结构和进化力量进行了全面的研究。值得注意的是,研究人员在 14 号染色体上发现了四个紧密连锁的色素沉积基因(BCO2和下游PTS),它们对雄性红色产生主要影响。此外,2 号染色体上的结构相关基因MAP7成为与雌性蓝色相关的潜在候选基因。研究人员的研究结果表明,类固醇激素相关基因(SULT2B1和SRD5A2)可能作为影响两性二态性着色的上游调控因子。此外,研究人员提出的证据表明这些颜色可能具有生物学意义。研究人员的研究结果表明,GN 中存在持续选择压,可能有助于红色和蓝色等位基因的固定,从而将 GN 表型限制在木格滩沙地。虽然这些观察结果很有趣,但研究人员建议未来的研究可以深入阐明这些位点的功能相关性和共同选择机制。此外,探索类固醇激素在性别二态性着色发展中的潜在作用可以为控制性别差异着色的调节过程提供有价值的见解。
相关阅读
-

在动物界,肤色的性双态变异是一种普遍现象,它对动物的生存和繁殖成功与否有着重要影响。然而,人们对这种变异的遗传基础仍然了解不足。...
-

人性人一半是野兽,一半是天使。由自然的眼光看,人是动物,人的身体来源于进化、遗传、繁殖,受本能支配,如同别的动物身体一样是欲望之物。由诗和宗教的眼光看,人是万物之灵...
-

初中化学微项目化学习课程的构建研究——以项目皮蛋的制作为例(化学教学2022年第1期)
摘要: 基于国家课程中的核心知识和关键能力,将项目化学习的设计要素融入课堂教学,有利于激发学生持续性的学习活...
-

Nature:遗传风险变异通过不同的途径导致 2 型糖尿病的发展
通讯作者单位:慕尼黑工业大学DOI:主要内容:迄今为止,最大的2型糖尿病全基因组关联研究,包括几个祖先群体,导致确定了八个遗传风险变异集群。...
-

一份属于每个人的生活简化指南...
-
江西医疗卫生群414420032︱公共基础知识每日一练︱医学专业知识每日一练泌尿系统由肾、输尿管、膀胱和尿道组成。其中肾为生成尿液的器官。在解剖学中...
-
初中化学微项目化学习课程的构建研究——以项目皮蛋的制作为例(化学教学2022年第1期)
摘要: 基于国家课程中的核心知识和关键能力,将项目化学习的设计要素融入课堂教学,有利于激发学生持续性的学习活...
-

如何用化学方法区分学霸与学渣?初中化学学习的三大难点在这儿!
总体来说,学霸和学渣以“混合物”的形式同时存在于学术的金字塔里。初中化学学习的三大难点。...
-
Video PlayerClose新闻中的即兴短评,一般是播音员或主持人依托新闻事件,以事实为导向,对新闻事件进行告知的同时...
-

点击文章领取学习资料...
-
辽宁招生考试之窗...
发表评论
版权声明:本文内容由互联网用户自发贡献,该文观点仅代表作者本人。本站仅提供信息存储空间服务,不拥有所有权,不承担相关法律责任如发现本站有涉嫌抄袭侵权/违法违规的内容,请发送邮件举报,一经查实,本站将立刻删除。
